

Fiziologia specială – profesor Gh. Petrescu
09.10.2002 Curs 2
Metabolismul miocardului:
Din sângele arterial O2 trece în lichidul interstiţial prin difuziune, iar în cardiomiocit trece tot prin difuziune. O2 se transformă în CO2 care este recoltat în venel coronare, iar apoi ajunge în atriul drept.
Arterele coronare au implicaţii fiziologice dar şi patologice.
Cardiomiocitul având nevoie de mult O2, arterele coronare trebuie să posede caracteristica de vasodilatatoare pentru a putea creşte fluxul sanguin.
=> histochimia fibrei miocardice:
» celule musculare albe:
o contracţie rapidă;
o mitocondrii în număr redus;
o enzime anaerobe;
» celule musculare roşii:
o contracţie lentă;
o mitocondrii numeroase;
o enzime aerobe;
¶ caracteristici metabolice:
· condiţii normale:
§ funcţionare în aerobioză;
§ saturaţie în O2 a sângelui venos scăzută;
§ mitocondrii dense;
§ enzimele glicolitice sunt puţin active;
§ catabolismul acizilor graşi crescut;
§ conţinut bogat în mioglobină – rezervor de O2;
· activitate contractilă redusă => sinteză redusă de: peptide, lipide, glucide;
¶ captarea substratului:
· organ aerob => captare de acizi graşi neesterificaţi;
· glucoză şi acid lactic => mai puţin decât muşchiul scheletic;
· substratul:
§ captat pasiv: A.G.N. şi Ac. A => gradient de concentraţie;
§ captat activ: glucoza <=> insulino-dependent;
¶ metabolismul glucidic:
· rezerve reduse de glicogen <=> catecolamine;
· glicoliza:
o anaerobă => ATP (singura sursă anaerobă)
o aerobă => ATP – ciclul Krebs (mitocondrii)
o lactic – dehidrogenaza:
§ miocardică (lactat => piruvat)
§ musculară (piruvat => lactat)
o fosfofructokinaza => fosforilarea hexozelor => catabolism
§ cantitate redusă
§ este reglată alosteric <=> ADP şi ATP <=> hipoxia scade ATP
În cadrul metabolismului glucidic avem 2 enzime principale: acid lactic (în celula miocardică lactatul se transformă în piruvat, iar acidul piruvic trece în ciclul Krebs).
O a doua enzimă care se găseşte în stare latentă este fosfofructokinaza, care are activitate redusă în condiţii obişnuite.
¶ metabolismul lipidic:
» acizi graşi => oxidaţi pe calea piruvatului (mitocondrie)
» acizi graşi => esterificaţi => gliceride (anoxie)
Acizii graşi au şi catabolism şi anabolism în cardiomiocite. În condiţii normale activarea acizilor graşi este caracterizatp de catabolismul lor.
În condiţiile în care acizii graşi în cardiomiocite nu pot fi activaţi ca atare, nefiind o cantitate de O2 necesară, acizii graşi sunt esterificaţi.
relaŢIA PRESIUNII o2 DIN SÂNGELE CORONARIAN ŞI PRODUCEREA DE ATP
□ cele două se condiţionează fiind în strânsă legătură
□ sângele venos coronarian ~ desaturat în O2
□ reţinerea de O2 de fibra miocardică => debit coronarian <=> adenozină
o => vasodilataţie coronariană
o <=> degradarea ATP
o prin travaliu intens – ATP
□ O2 => mioglobină => mitocondrii => ATP
o contracţie
o PC (fosfofructokinază)
Sursa de energie necesară este asigurată de ATP. Adenozina produce vasodilataţie.
¶ Metabolismul protidic:
· rol energetic <=> puţin utilizat
· sinteza de proteine:
o mecanism obişnuit => ARNm => ribozomi
o poliamidele (spermina, putresceina) => stimularea sintezei proteice => hipertrofie cardiacă => ATP => (inhibă) => ornitindecarboxilaza => poliamide
· sinteza proteinelor contractile:
o miozina:
§ se sintetizează mai lent faţă de proteinele citoplasmatice
§ mai rapid decât miozina musculară scheletică
o sediu – mitocondrie (ADN mitocondrial)
· sinteza ARN (în nucleu):
o riboza:
§ miocard (ciclul pentozelor)
§ acizi aminaţi + fosforilare oxidativă
o lipsa mitozelor în miocard (diviziune amitotică)
o hipertrofia miocardică => ARN ribozomal => captarea amino-acizilor => creşte masa miocardului
Poliamidele stimulează sinteza de proteine. Scade cantitatea de ATP când există disconcordanţă între proteine şi necesităţi. Mioglobina este o enzimă sarcoplasmatică. Mitocondriile se găsesc abundent în cardiomiocite.
Metabolismul ATP-ului şi ca2+ la nivelul miocardului
Rolul ATP miocardic:
» sinteza de proteine
» 90% – funcţia proteinelor contractile
» 10% – fenomene de transport activ
ATP-aze:
» membranare
» reticul sarcoplasmatic
» mitocondrie – sistem complex
» sarcomer
Potenţial de repaus: – ATP-aza Mg2+:
□ stimulată de Na+ şi K+
□ inhibată de digitalice – strofantina (cobaina)
Ca2+:
□ ATP-aza Ca membranară => Ca2+ intracelular => platoul potenţialului de acţiune
□ intrasarcoplasmatic:
o puţin = 10-7 M
o vezicule => ATP-aza Ca dependentă
□ deplasarea Ca2+ => interior (platou potenţial de acţiune) => sarcomer (activat) => cantitate mică reticul sarcoplasmatic
metabolism cardiac în cursul efortului şi hipoxiei
Efectele supraîncărcării mecanice:
1) creşterea consumului energetic
a. creşterea consumului de O2 <=> vasodilataţie coronariană => creşte debitul coronarian
b. creşte consumul de substrat – acizi graşi neesterificaţi
2) stimularea sintezei de proteine
» apariţia poliribozomilor (½ h de anoxie)
» încorporare marcată de acizi aminaţi în miofibrile
» insuficienţă cardiacă => sinteză de proteine anormale
Efectele hipoxiei:
□ necesitatea de O2 pentru contracţia miocardică
□ consecinţele lipsei de oxigen:
1. scăderea ATPde sursă aerobă:
» producţie anaerobă de ATP (efect Pasteur)
» modificări ale transortului de ioni prin membrană (potenţial de membrană)
2. creşterea fosfor anorganic, ADP, adenozină:
» stimularea fosfofructominazei (degradare anaerobă)
» creşte captarea de glucoză
» acumulare:
o 1 glicerofosfat => trigliceride
o piruvat => lactat => scade pH-ul
» stimularea glicogenolizei
» formare adenozină => vasodilataţie coronariană
3. blocarea fosforilărilor oxidative:
» acumulare de vacuole de lipide
» acumulare H+ => creşte permeabilitatea membranară => eliminare în circulaţia generală a: H+, K+, fosfat, enzime (lacticdehidrogenaza)
4. stimularea eliberării de catecolamine <=> anoxie
5. blocarea sintezei de proteine:
Hipoxia: – incapacitatea cardiomiocitelor, de a-şi sintetiza ATP în condiţii aerobe.
Glucoza este asigurată din rezervele de glicogen (care sunt cam puţine, precare). Adenozina încearcă să aducă O2 necesar pentru travaliu–acumulare în vacuole.
Factorul care contribuie la astfel de situaţii în miocard este aportul de O2.
Proprietăţile muşchiului cardiac:
□ excitabilitate
□ automatism
□ ritmicitatea
□ conductabilitatea
□ contractilitatea
□ troficitatea
excitabilitatea miocardică
Definiţie: – proprietatea cardiomiocitului de a genera potenţial de acţiune la acţiunea unui stimul.
Cardiomiocitul când este excitat se prezintă în stare depolarizată. Excitantul (mai ales despre cel ventricular adult) normal, fiziologic al miocardului este potenţialul de acţiune generat în nodulul sino-atrial. Chiar în condiţii fiziologice inima poate să reacţioneze, să se excite datorită potenţialelor de acţiune pornite din diferite părţi ale miocardului.
În condiţii anormale inima se poate activa, contracta, excita datorită stimulilor din alte zone numite focare ectopice (care pot apărea oriunde în inimă, datorită unor influenţe extrinseci).
Este o celulă zisă–excitabilă (reacţionează la stimuli prin fluxuri ionice deosebite–generatoare de alt potenţial) Þ ionii trec prin membrană prin transport pasiv, una din ele fiind canalele ionice.
Principalele canale ionice (voltaj-dependente) din membrana cardiomiocitului concură la realizarea potenţialului de acţiune (asigură fluxuri ionice ce determină potenţial de acţiune) => ex: canale de Na+ rapide, voltaj-dependente (cu cele 2 bariere: m şi h).
Filtru de selectivitate = bariera m = fosfolipide membranare încărcate electric pozitiv, care în condiţiile activităţii stimulilor se reorientează în membrană. Bariera se deschide timp de 1 milisecundă, după care revine şi reapare bariera.
Canale de K+ lente:
Au implicaţii în depolarizare, dar mai ales în repolarizare membranară. Canalele de K+ de tip X1 care se deschid din momentul în care la 1 ms de la declanşarea potenţialului de acţiune ating maximul şi apoi se închid.
Canalele lente de K+ de tip K1 care asigură afluxul lent din repaus al ionului respectiv. Acest tip de canale este responsabil de generarea potenţialului membranar respectiv.
Acest tip de canale este responsabil de generarea potenţialului membranar de repaus.
Canale de Na+-Ca2+ lente:
Aceste canale asigură influxul de ioni de Ca+ şi Na+ generator de potenţial de acţiune.
Canale de Cl– rapide:
Vârful potenţialului de acţiune este rezultatul fluxurilor ionice de Cl (rapide, voltaj-dependente => când potenţialul membranar atinge valoarea 0).
Potenţialul de repaus:
» -80-85 mV
» pentru transport activ
» ce atinge echilibrul de K+
În condiţii normale cardiomiocitul adult este excitat de potenţialele de acţiune generate în nodulul sino-atrial.
Potenţialul de acţiune a cardiomiocitului adult diferă net de cel a neuronului sau a cardiomiocitului scheletic structural. Se modifică permeabilitatea ionică, influx masiv de ioni.
În faza 0 se ajunge la +30 mV de la -80-85 mV. Potenţialul de acţiune a cardiomiocitului adult durează 150-300 ms.
Potenţialul de acţiune:
¶ fazele depolarizării:
o platou:
§ 0,15-0,30 secunde
§ canale de Ca2+ şi Na+
§ permeabilitate K+ <=> scăzută
§ perioadă refractară lungă – tetanizarea nu este posibilă
o faza descendentă:
§ Na+ – canale rapide şi potenţialul de acţiune = 90-105 mV
o durata potenţialului de acţiune:
§ nerv – 3-4 milisecunde
§ fibră miocardică ~ 300 milisecunde
o prag critic:
§ 70 mV
¶ repolarizare – vârful potenţialul de acţiune are tendinţă efemeră de repolarizare a miocardului.
o repolarizare rapidă:
§ activare canale de Cl–
§ inactivarea canalelor de Na+ rapide
§ impermeabilitate pentru Na+
· => difuzie
· mecanisme membranare active => potenţial de repaus
o repolarizare moderat „rapidă”
§ inactivarea Na+, Ca2+
§ deschiderea celor de K+
§ se ajunge la potenţial de repaus
Dinamica canalelor ionice din miocard dă un aspect particular, celula se comportă diferit la acţiunea unor stimuli.
De ce inima nu se tetanizează? Din punct de vedere al contracţiei este o celulă care se contractă lent.
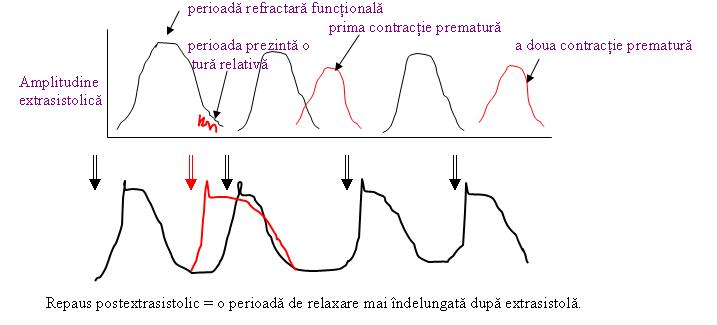
Perioada refractară a miocardului:
» inaxcitibilitate => potenţial de acţiune – „perioada refractară absolută”
» excitant:
o sfârşitul platoului => potenţial nepropagabil
o stimul normal
» perioada refractivă funcţională:
o auricular = 0,15 secunde
o ventricul = 0,25-0,30 secunde => primul răspuns propagabil
» perioada refractivă relativă:
o auricular = 0,03 secunde
o ventricular = 0,05 secunde
» contracţie precoce:
o extrasistolă
o repaus postextrasistolic
o ineexcitabilitate periodică
Inima este inexcitabilă o lungă perioadă de timp 300 milisecunde, dacă un stimul acţiunea asupra inimii, cardiomiocitul nu răspunde deloc indiferent de intensitatea stimului = perioadă refractară absolută.
La sfârşitul platoului, doar un stimul cu intensitate mare determină un potenţial, dar care nu se poate propaga. Acelaşi stimul cu intensitate mare către sfârşitul pantei de polarizare şi produce un potenţial de acţiune propagabil.
automatismul şi ritmicitatea miocardului
Automatism:
» proprietatea miocardului de a genera potenţial de acţiune
» lipsa influenţelor extrinseci
Toate celulele miocardice au automatism. Celulele miocardice adulte au această capacitate lentă. Capacitatea miocardului de autogenerare a potenţialului de acţiune (de descărcare spontană a potenţialului de acţiune).
Mecanism de producere:
¶ funcţia miocardului adult => potenţialde repaus: -80-90 mV şi este stabil
¶ fibra miocardului embrionar:
o potenţial de repaos –60-70 mv – instabil / potenţial maxim diastolic
o depolarizare lentă diastolică ~ 55 mV
¶ desfăşurare:
o potenţial repaos:
§ scăzut
§ permeabilitate pentru Na+ şi K+
o sfârşitul potenţialului de acţiune:
§ g K+ Þ crescută şi g Na+ => foarte scăzută
§ ecuţia Goldman = g K+/g Na+ => crescută (potenţial de membrană negativ)
§ K+ extracelular Þ hiperpolarizare
o potenţial maxim diastolic:
§ scăderea g K+
§ zecimi de secundă Þ Na+ lent
· => celulă
· => prag de excitabilitate => potenţial de acţiune
